the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 17 Sep 2025
| 17 Sep 2025
Carbon–climate feedback higher when assuming Michaelis–Menten kinetics of respiration
Christian Beer
Earth system models simplify complex terrestrial respiration processes assuming a first-order chemical reaction or assuming a Michaelis–Menten kinetics. The effect of the respective mathematical representation on the terrestrial carbon–climate feedback is unclear. Using a simplified model of biogeochemical feedbacks to climate, I show that the terrestrial carbon–climate feedback roughly doubles when assuming Michaelis–Menten kinetics of respiration. Consequently, the remaining carbon budget to keep global warming below 2 °C is substantially higher. The effects of the respiration formulation also depend on the underlying emission scenario. These results highlight the importance of an increased understanding of the respiration processes on a global scale to more reliably project future carbon dynamics and climate and related feedback mechanisms and thus to estimate a valid remaining anthropogenic carbon budget using Earth system models.
- Article
(1192 KB) - Full-text XML
-
Supplement
(343 KB) - BibTeX
- EndNote




The anthropogenic emission of carbon dioxide into the atmosphere since the industrialization period has led to a global warming of about 1 K due to the greenhouse effect (Canadell et al., 2023). However, less than half of the anthropogenically emitted carbon remains in the atmosphere because terrestrial ecosystems and the ocean take up 34 % and 25 %, respectively (Friedlingstein et al., 2023). The main reasons for this strong carbon dioxide uptake in terrestrial ecosystems are biogeochemical feedbacks (Cox et al., 2000). Increasing atmospheric carbon dioxide (CO2) concentration leads to an enhanced photosynthesis rate and hence to CO2 uptake by vegetation on land (Cramer et al., 2001; O'Sullivan et al., 2022). This carbon is stored in vegetation pools and ultimately transferred to soils by exudation, litterfall, and mortality processes, thereby increasing the soil carbon content. his is the important carbon-concentration feedback mechanism (Arneth et al., 2010) (Fig. 1), which is a negative feedback and hence responsible for the current net CO2 sink on land that has been preventing us from even stronger climate change. In contrast, autotrophic respiration and heterotrophic respiration are also higher than under pre-industrial conditions (Canadell et al., 2023) due to (i) higher substrate availability and (ii) the positive temperature sensitivity of respiration (Lloyd and Taylor, 1994). This temperature sensitivity of respiration forms the positive carbon–climate feedback mechanism (Fig. 1): higher CO2 concentration leads to higher temperature, which increases respiration and hence leads to an even higher atmospheric CO2 concentration (Arneth et al., 2010).
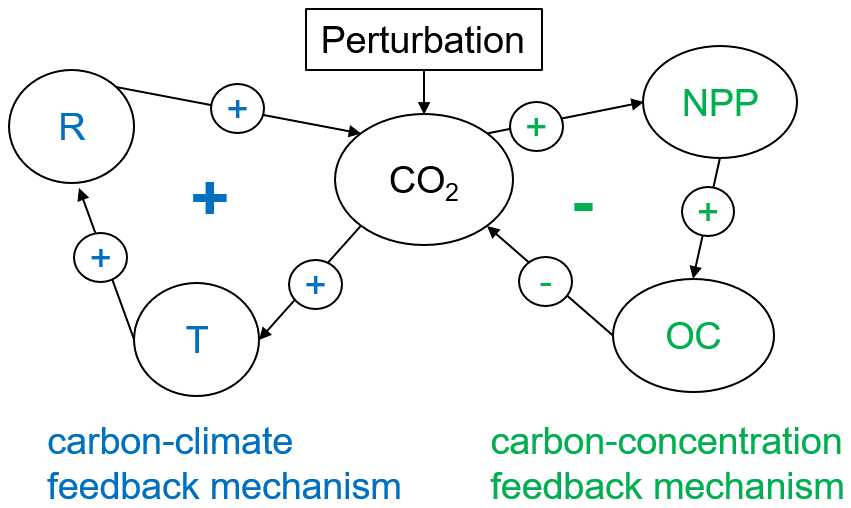
Figure 1Feedback diagram for two main terrestrial biogeochemical feedback mechanisms. NPP: net primary production, R: respiration, OC: land organic carbon stocks, T: global surface air temperature, CO2: atmospheric carbon dioxide content.
These two biogeochemical feedback mechanisms have been identified as two major feedback mechanisms in the Earth system, with great impact on climate (Friedlingstein et al., 2006; Arora et al., 2020). Currently, the positive carbon–climate feedback is lower than the negative carbon concentration feedback, and therefore land ecosystems act as a natural sink of CO2 of about 3 Pg C yr−1 (Friedlingstein et al., 2023). However, due to internal dynamics of the system, climate change, and changes in anthropogenic CO2 emissions, the future strength of the feedback mechanisms, and hence the net CO2 exchange between land and atmosphere, remains unclear. Recent accumulation of soil carbon in concert with higher future temperature and a declining increase in productivity can lead to a decreasing land sink under increasing CO2 emissions in future (Jones et al., 2023; Cramer et al., 2001). To estimate such feedbacks, we need to run a modified version of an Earth system model in which only one feedback mechanism is considered. The temporal difference in atmospheric CO2 concentration from such experiments to model runs without the feedback is used to quantify these feedbacks (Hansen et al., 1984).
For the carbon–climate feedback mechanism (Fig. 1), the representation of respiration processes in Earth system models is crucial. Several assumptions about the underlying processes and respective mathematical representations have been proposed. Land surface models usually represent respiration as a linear function (first-order kinetics) to the amount of available substrate (organic carbon, C),
using several carbon pools (Sitch et al., 2003; Brovkin et al., 2013; Tang et al., 2022), with different decomposition rate constants k. In doing so, we assume that the active microbial biomass pool increases in relation to increased substrate availability. However, the underlying biochemical reactions are mostly enzymatic; hence a Michaelis–Menten kinetics model has been proposed to represent the dynamics of respiration (Wieder et al., 2013; Yu et al., 2020):
where vmax is the maximum reaction rate under infinite carbon substrate C, and KM represents the amount of carbon at which the reaction rate is half of the maximum. In this model, we assume a constant active microbial biomass pool. The non-linear shape of this relationship between reaction rate and substrate availability (in contrast to the linear dependency of first-order kinetics models) leads to a steep increase of the reaction rate under low substrate availability while only a moderate to negligible increase under high substrate availability. In doing so, this model implicitly represents the function of enzymes in the underlying biochemical reactions. It has been proposed that such a model enables a more valid aggregation from the process level (e.g. rhizosphere, aggregatusphere) to the landscape scale (Reichstein and Beer, 2008). However, both models are great simplifications of the underlying biogeochemical processes with strong assumptions. Therefore, in this study I will use both equations to represent respiration and study the resulting structural uncertainty in feedbacks and remaining C budgets.
The two approaches represented by Eqs. (1) and (2) imply different responses of respiration to changing substrate availability. Therefore, future dynamics of respiration should differ depending on the mathematical formulation. Such structural model uncertainty is of interest in particular because there might be a point when the land sink starts to decrease even under continuing high anthropogenic emissions (Cramer et al., 2001) but also because of the question of how land sinks will react to decreasing or even negative anthropogenic carbon emissions.
Therefore, I ask three main questions in this paper: what is the effect of the respiration model structure on
-
projections of the land carbon sink,
-
the strength of the carbon–climate feedback and
-
the remaining anthropogenic carbon budget
under different carbon emission scenarios? To address these questions I performed a feedback analysis using a simplified but process-based model of global biogeochemical feedback mechanisms twice, using a first-order and a Michaelis–Menten kinetics model of respiration. The simplified model of global biogeochemical feedback mechanisms is of zero dimension (only globally aggregated pools) and neglects many detailed processes and interactions between ecosystem components. Therefore, the idea is not to precisely quantify C budgets or feedback but rather to show the effects of the respiration model structure on these estimates qualitatively.
2.1 Simplified carbon–climate feedback model
The model has been designed to study the two major biogeochemical feedbacks to climate displayed in Fig. 1. Exchanges of carbon among atmosphere, ocean, and land are represented using a reduced number of carbon pools without spatial details but still in a process-based way, i.e. based on a set of differential equations. For example, the amount of carbon taken up by vegetation depends on the atmospheric carbon content, while the amount of CO2 released to the atmosphere due to respiration depends on the carbon content of the ecosystem. The model assumes a global surface air temperature response to changing atmospheric carbon dioxide content using a transient climate response parameter, which is lagged due to the ocean heat capacity. The model is driven by anthropogenic carbon dioxide emissions to the atmosphere following several scenarios developed for the IPCC 6th assessment report.
A detailed description of the model can be found in Lade et al. (2018). Here, I apply two alternative model versions, one assuming a first-order kinetics of respiration (FOK) and one assuming a Michaelis–Menten kinetics of respiration (MMK). The representation of terrestrial carbon uptake by gross primary productivity (GPP) is identical in both model versions. It is assumed to increase logarithmically with atmospheric carbon dioxide Ca (Eqs. 3 and 4, first term right-hand side). In addition, emissions due to land-use change EL are subtracted the same way in both versions, and the increase in respiration with temperature is represented by a typical Q10 model (Eqs. 3 and 4, second term right-hand side). Only the dependence of respiration to land carbon stocks differs. The FOK model assumes a first-order kinetics with a respiration rate constant estimated by pre-industrial GPP and carbon stocks, , following the same principle as in Lade et al. (2018).
In contrast, the MMK model represents respiration as a classical Michaelis–Menten equation with parameters vmax and KM:
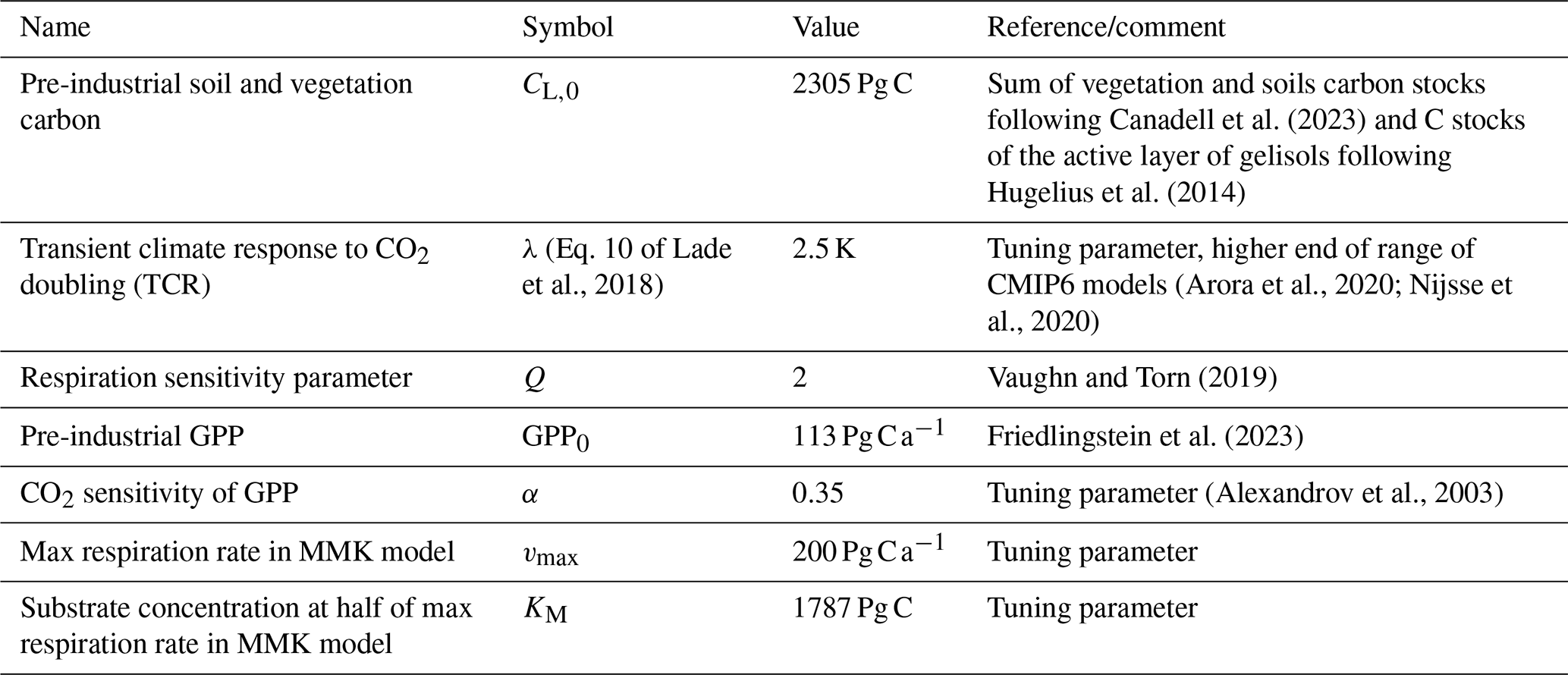
Parameters and pre-industrial pools and fluxes for model initialization were taken from Lade et al. (2018) and partly adjusted (Table 1). The transient climate response to CO2 doubling λ is set at the higher end of the range reported for CMIP6 model results (Arora et al., 2020) in order to match the observed historical temperature anomaly. Parameters vmax and KM of Eq. (4) are optimized using a standard gradient decent approach (MATLAB R 2023b function lsqnonlin) such that the difference of the modelled and observation-based land carbon changes is minimized.
Table 1Value and description of parameters different from Lade et al. (2018).
2.2 Modelling protocol
The two model versions have been run from 1850 until 2100 using a daily time step forced by anthropogenic carbon dioxide emissions from fossil fuel burning and from land-use change. For this, I combined reported historical emissions from the Global Carbon Project (Friedlingstein et al., 2023) with Shared Socioeconomic Pathway (SSP) emission scenarios from the public database of the Institute for Applied Systems Analysis (Riahi et al., 2017). I selected four widely used scenarios produced for the CMIP6 protocol (Gidden et al., 2019): SSP1-26 (optimistic scenario, reaching economic growth while retaining sustainability and reducing inequalities), SSP2-45 (including mitigation strategies), SSP3-70 (represents a future of inequality and fossil fuel dependency), and SSP5-85 (representing economic growth through strong reliance on fossil fuels). These scenarios reach a forcing of 2.6, 4.5, 7.0, and 8.5 W m−2 at the end of the century and represent a huge spread of carbon emissions into the atmosphere (Fig. 2). I linearly interpolated the reported emissions at decadal scale to an annual resolution. In the combined time series (Fig. 2), historical emissions span the period 1850–2014, and scenarios continue from 2015 until 2100.
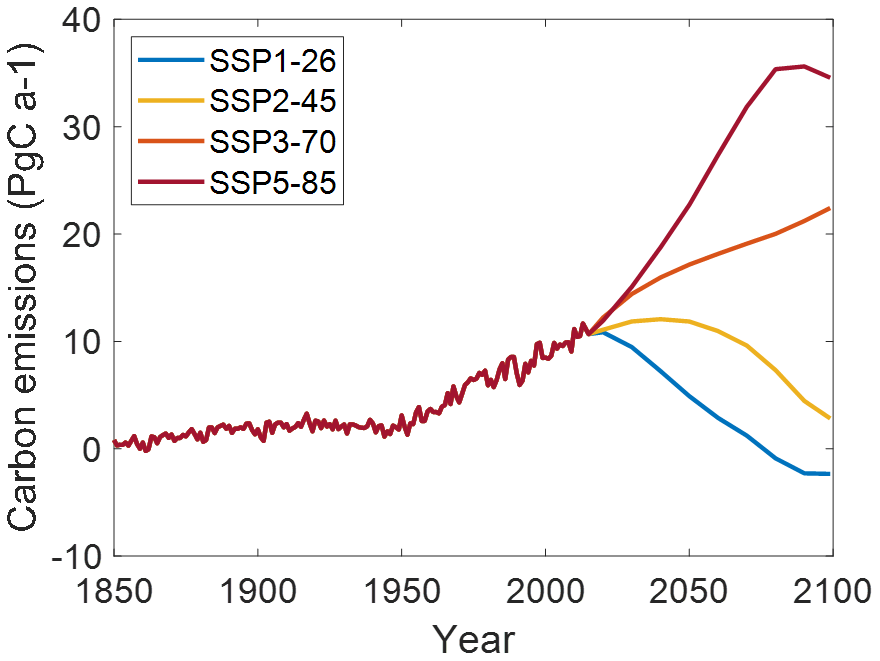
Figure 2Total CO2 emissions from burning fossil fuels and land-use change from combining a historical dataset with results from Integrated Assessment Models for different scenarios.
I performed model simulations for these emission scenarios and for both model versions, FOK and MMK. The results were used to evaluate the model during the historical period and to estimate the remaining carbon budgets to keep warming below a certain threshold. For the feedback analysis, all these simulations were repeated three times. To estimate the feedback factor, I did model simulations in which only the terrestrial carbon–climate feedback is considered. The results were used to estimate the respective (Sect. 2.4). For calculating the feedback sensitivities β and γ (Sect. 2.4), I additionally performed biogeochemically and radiatively coupled simulations following Friedlingstein et al. (2006) and Lade et al. (2018) and derived ΔCL, ΔCA, and ΔT from these simulations. In the biogeochemically coupled simulation, I set λ to 0; hence effects of CO2 change on temperature are excluded. In the radiatively coupled simulation, I neglected all effects of CO2 on terrestrial or marine carbon pools. In total, there are 32 model simulations.
2.3 Feedback analysis
Atmospheric carbon content increases in time due to annual anthropogenic emissions (ei) and internal feedback mechanisms. To estimate this carbon dioxide change when considering a terrestrial carbon–climate feedback (“on”), I averaged the atmospheric carbon content during a reference period in the future (2080–2100) and in the past (1850–1900) using the respective model simulation (Sect. 2.3) and subtract both:
The respective atmospheric carbon change without considering the feedback (“off”) equals the sum of emissions:
The feedback is the difference , and the feedback factor F is the ratio of both changes, which can be used to compare feedbacks and to identify positive (F>1) or negative feedbacks (F<1) (Cox et al., 2000; Friedlingstein et al., 2003; Hansen et al., 1984; Zickfeld et al., 2011):
Sensitivities of the land carbon change to atmospheric carbon concentration (β) and temperature changes (γ) are defined following Friedlingstein et al. (2006) and Heinze et al. (2019) as
I used the biogeochemically coupled simulation results to estimate β (ΔT=0) and the radiatively coupled results to estimate γ (ΔCA=0).
Model results of carbon fluxes and the surface temperature anomaly for the historical period are in general agreement with results by the Global Carbon Project (Friedlingstein et al., 2023) and the NOAA Global Surface Temperature record (Fig. 3); i.e. the overall historical trends are captured. The model does not represent spatial details; oversimplifies functional diversity; and does not represent certain processes, such as disturbances. Therefore, the model is not able to capture the inter-annual variability of land carbon fluxes (Fig. 3). The general long-term agreement shows that major biogeochemical feedback mechanisms are correctly represented and that initial conditions (Table 1) and model parameters (Table 1) are reasonable. Therefore, we assume that we can apply this model to study the effects of structural respiration model uncertainty on the carbon–climate feedback strength.
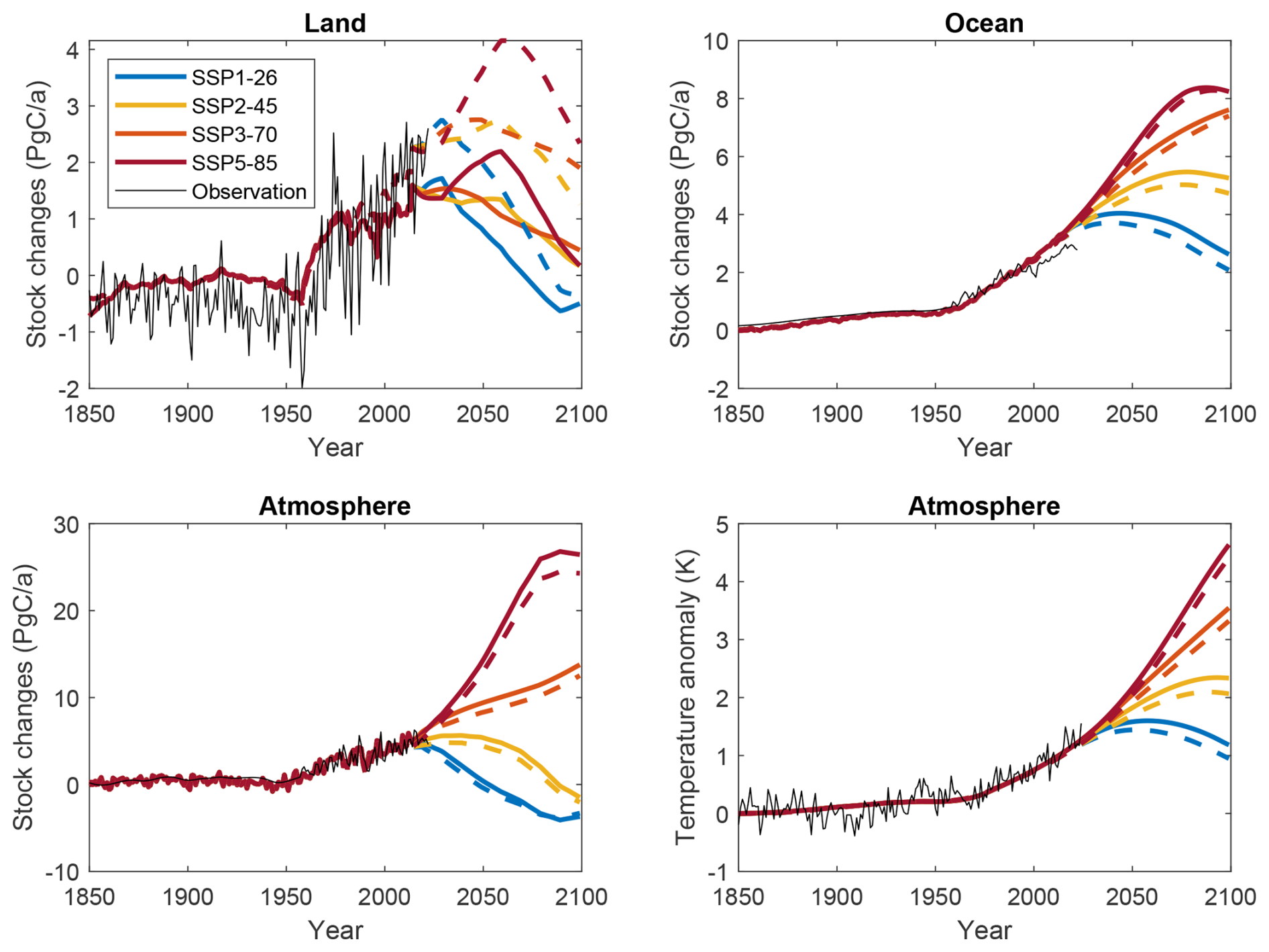
Figure 3Simulated carbon fluxes and temperature anomaly for the different scenarios. FOK and MMK model results are displayed by solid and dashed lines, respectively. Simulation results are compared to estimates by the Global Carbon Project or to the NOAA Global Surface Temperature record, which has been bias-corrected to the model results to match reference periods.
Figure 3 also shows the projections of carbon fluxes to land, ocean, and atmosphere, as well as the temperature change for the two different model structures until 2100 following the different emission scenarios. Overall, these projections of the main carbon cycle fluxes and temperature change are similar to concentration-driven CMIP6 results (Canadell et al., 2023; Jones et al., 2023). The projected ocean carbon sink in this study is substantially higher for most of the scenarios, and the land carbon sink of the model using a first-order kinetics respiration approach (FOK) is lower but comparable with Lade et al. (2018). Otherwise, the projections of the change in atmospheric carbon stocks and the global surface temperature change are similar to studies using Earth system models (Canadell et al., 2023). However, spread of carbon cycle projections using other models is usually also very high (Canadell et al., 2023; Jones et al., 2023), and the uncertainty due to parameter values or initial conditions hardly quantified in these studies.
The projected land sink evolution differs depending on both the emission scenario and the model structure applied. Under high-emission scenarios, the land sink continues to rise and peaks in the middle of the century followed by a decreasing sink until 2100. This peak has been reported by dynamic global vegetation models (DGVMs) and Earth system models (ESMs) before (Cramer et al., 2001; Jones et al., 2023) and is due to the reverse shape of the two main response functions, logarithmic productivity response to elevated CO2 and quasi-exponential respiration response to temperature. A second reason is internal carbon dynamics: respiration depends on the amount of land carbon stocks, which continued to increase until some maximum and therefore is the basis for a high respiration flux during the following time. For the scenario SSP1-26, the land sink starts to decrease immediately after the historical period, i.e. when emissions are reduced, and, depending on the model structure, is even becoming negative in the second half of the century.
The projected land carbon sink in 2100 is much higher when assuming a Michaelis–Menten kinetics model for respiration (MMK) even under an equal temperature sensitivity of respiration as by the first-order kinetics model (FOK), and even when parameters are chosen to fit both model results during the historical period. In addition, the peak in the middle of the century is more pronounced when using the MMK model (Fig. 3). Hence, this difference is only due to internal carbon dynamics differences, in particular a non-linear (decreasing) change of the respiration rate with increasing substrate availability when assuming Michaelis–Menten kinetics. This clearly demonstrates the uncertainty of land carbon sink dynamics just due to alternative assumptions and mathematical formulations of respiration processes. As a result of higher land sinks using the MMK model, ocean and atmosphere sinks are smaller, and the temperature change is lower (Fig. 3). Due to the higher land C sink assuming Michaelis–Menten kinetics, total changes in land carbon stocks are also much higher; i.e. land takes up several hundreds of Pg C more depending on the emission scenario.
These differences in the projected land sinks do have clear consequences for the transient climate response to cumulative emissions of carbon dioxide (TCRE) and hence the remaining anthropogenic carbon budgets under different emission scenarios. Usually, there is a quasi-linear relationship between the cumulative emission and the temperature change (Fig. 4). Under reduced emissions of SSP1-26 scenario, ocean and land C uptake may remain high (blue curves in Fig. 3), leading to a hysteresis in the TCRE (Koven et al., 2023). Such hysteresis is not visible in the other scenarios (Fig. 4) because emission reductions are not strong enough (Fig. 2). Interestingly, the relationship is less steep and more non-linear for the MMK model for all scenarios. From the TCRE the remaining carbon budget for a certain temperature threshold can be estimated (Canadell et al., 2023). In Fig. 4, the vertical lines indicate the amount of emissions since 2024 that – according to this model – can still be emitted in order to keep warming below the threshold of 2 °C warming compared to the pre-industrial situation, which is indicated by the horizontal line. We skip this analysis for scenario SSP1-26 results because the MMK model fails to reach a 2 °C increase at all (Fig. 4). For the other emission scenarios, the FOK model suggests 381 to 423 Pg C that can be emitted to the atmosphere in order to keep warming below 2 °C compared to pre-industrial temperature (Fig. 4). These estimates are slightly higher than the median remaining C budget estimated by CMIP6 experiments using ESMs of 370 Pg C (Table 5.8, Canadell et al., 2023). Importantly, when assuming a Michaelis–Menten kinetics of respiration (MMK), the remaining C budget is higher and ranges between 457–536 Pg C. This is due to flatter slopes of these model results (Fig. 4).
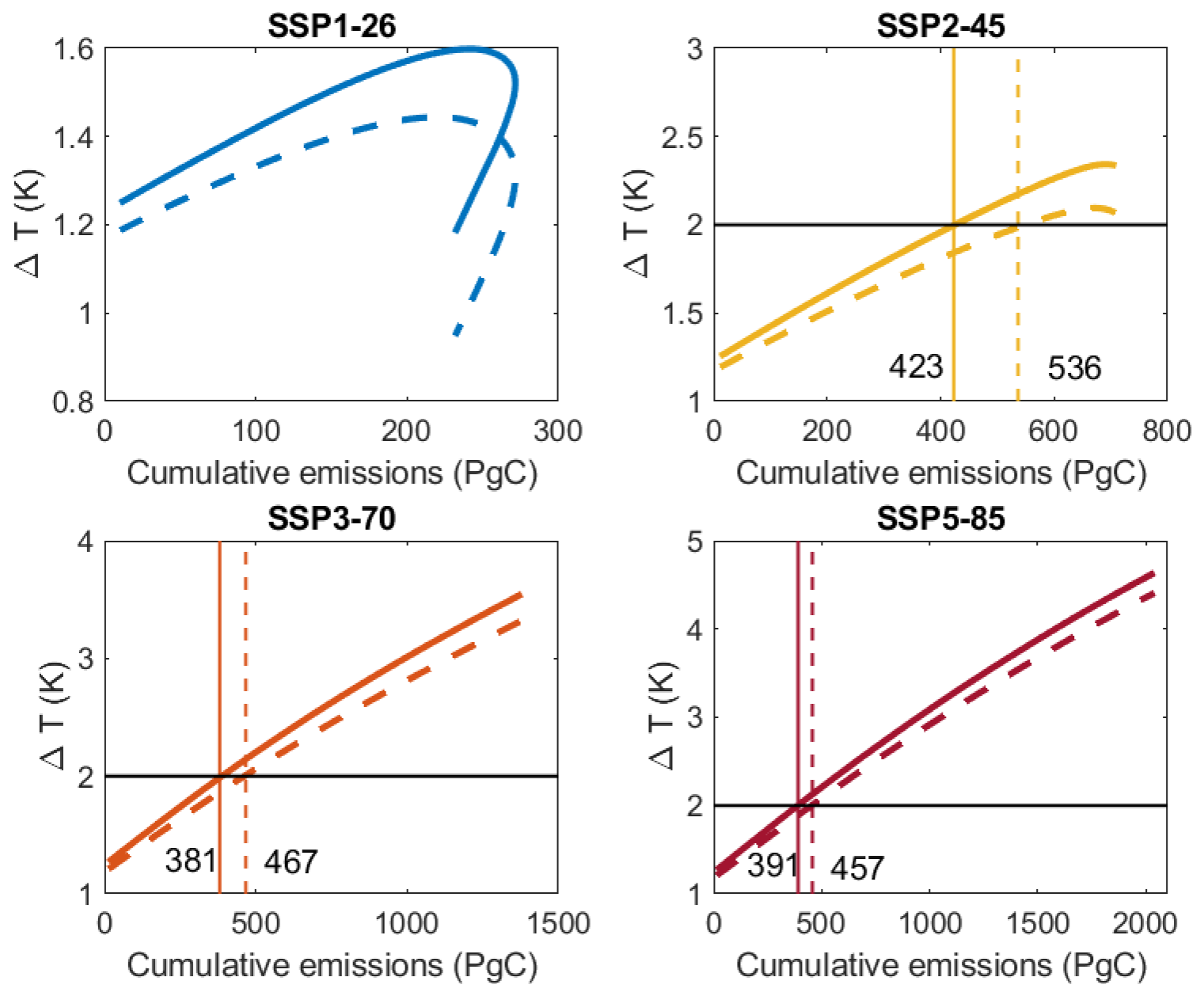
Figure 4Relationship between global air surface temperature difference to pre-industrial temperature and the cumulative emission of CO2 from 2024 until 2099 for different emission scenarios and the two model simulations FOK (solid lines) and MMK (dashed lines). Horizontal lines indicate a temperature change threshold of 2 K, and vertical lines and numbers indicate the respective cumulative emissions since 2024 to reach that temperature change target.
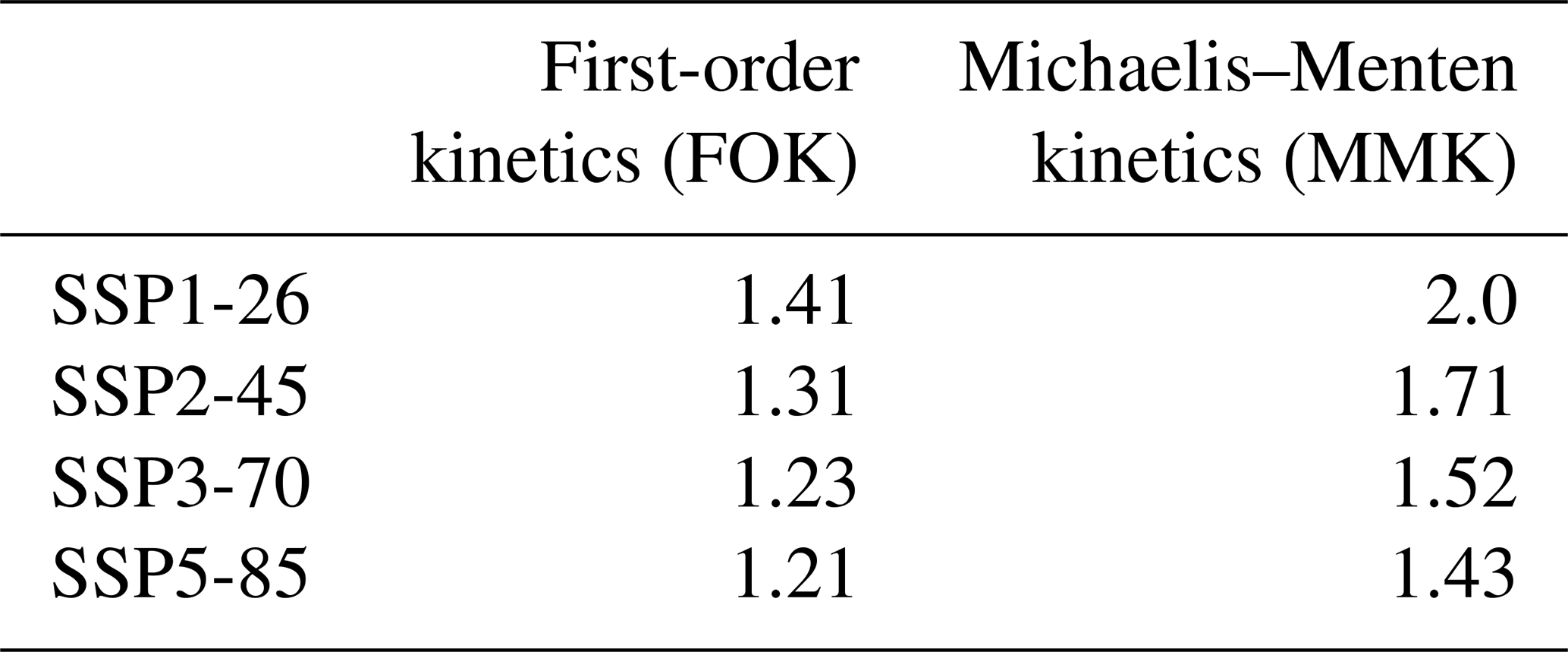
Using the first-order kinetics approach of respiration (FOK), I estimate a carbon–climate feedback of 379 to 498 Pg C when comparing the average CO2 concentration of the period 2080–2100 with pre-industrial conditions, depending on the emission scenario (Table 2). This translates into feedback factors of 1.2 to 1.4 (Table 3), which are similar to previous estimates (Lade et al., 2018). Interestingly, the strength of the feedback mechanism as expressed by the feedback factor decreases with increasing carbon emissions (Table 3); i.e. the internal Earth system interactions are more important under reduced anthropogenic emissions. However, when assuming Michaelis–Menten kinetics of respiration, this carbon–climate feedback strength is higher (Table 3) depending on the underlying scenario.
Table 2Terrestrial carbon–climate feedback (Pg C) for different representations of respiration in the model. Shown is the difference of model results accounting for the feedback and excluding it based on the temporal change in atmospheric carbon content between 2080–2100 and 1850–1900.
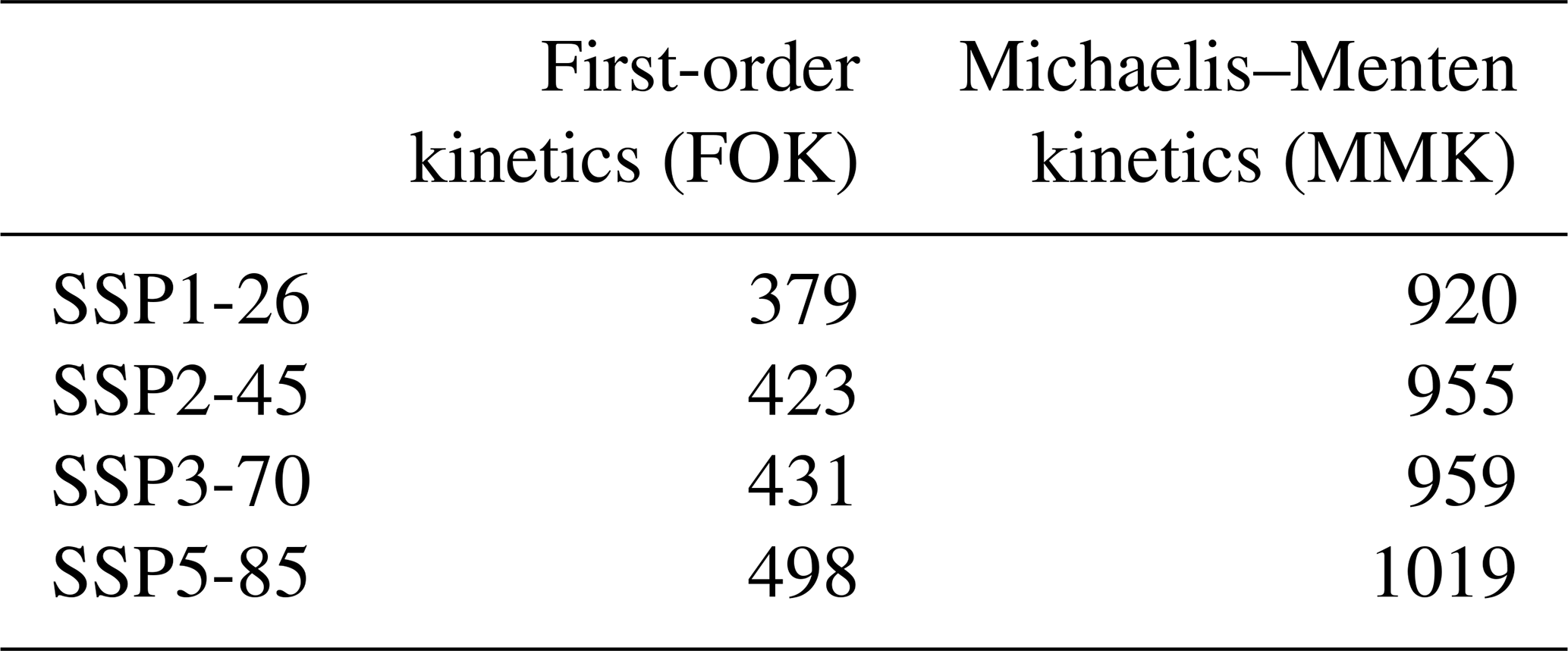
Table 3Feedback factor of the terrestrial carbon–climate feedback for different representations of respiration in the model. Shown is the difference of model results accounting for the feedback and excluding it based on the temporal change in atmospheric carbon content between 2080–2100 and 1850–1900.
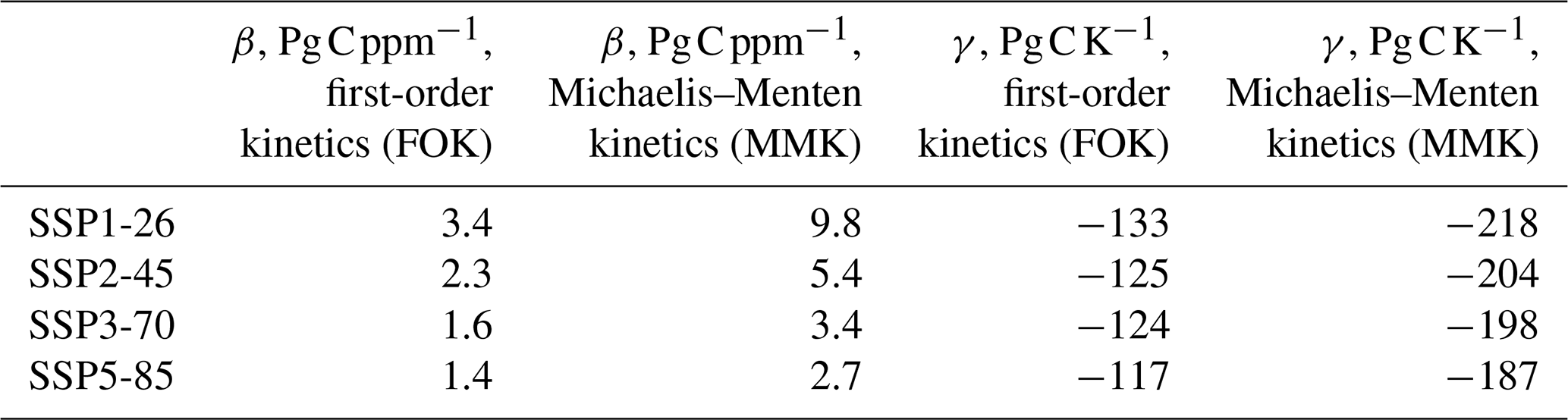
The FOK model estimates the sensitivity of the land carbon change to increasing atmospheric CO2 concentration (β, Table 4) to be 1.4 Pg C ppm−1 assuming the high-emission scenario SSP5-8.5. This is similar to CMIP4 model runs using the high-emission scenario SREAS A2 (Friedlingstein et al., 2006) and at the higher end of the range of CMIP6 model results without considering the N cycle in 4×CO2 experiments (Arora et al., 2020). Interestingly, the sensitivity increases towards scenarios assuming fewer emissions (Table 4), and the sensitivity is higher when assuming a Michaelis–Menten kinetics of respiration (Table 4). The land carbon change sensitivity to climate change (γ, Table 4) is estimated at −117 Pg C K−1 in this case. This is at the higher end of the range for the previously mentioned ESM results (Arora et al., 2020; Friedlingstein et al., 2006). This parameter is also more negative when assuming Michaelis–Menten kinetics or when considering a lower emission scenario (Table 4).
Table 4Sensitivities of the land carbon change to changing atmospheric carbon dioxide (β, Pg C ppm−1) and temperature (γ, Pg C K−1) for different representations of respiration in the model (FOK and MMK).
Besides gross primary productivity, ecosystem respiration is one of the main land–atmosphere carbon exchange processes (Friedlingstein et al., 2023). The underlying biochemical processes are complex, and mathematical models of simplified net reactions are usually applied in Earth system models: either assuming a first-order chemical reaction of carbon and oxygen to carbon dioxide and applying Eq. (1) or considering the underlying enzymatic reactions and hence applying Eq. (2). The epistemic uncertainty in projecting future land–atmosphere exchange of CO2, climate, and the related biogeochemical feedbacks underlying these assumptions has been addressed in this paper. Model parameters have been chosen based on literature values and to fit published historical carbon and temperature changes (Sect. 2.3) for the first-order kinetics approach (FOK).
For the Michaelis–Menten kinetics model (MMK), we selected parameter values such that results are also similar to Global Carbon Budget estimates and the FOK model during the pre-industrial period. Interestingly, effects of anthropogenic carbon emissions on future land sink dynamics differ between both model versions, with several Pg C yr−1 higher uptake by land when assuming Michaelis–Menten kinetics for respiration (Fig. 3). Such higher land carbon uptake leads to a lower ocean carbon sink, hence increasing differences between land and ocean sinks. In addition, the projected global surface temperature change until 2100 is lower in the MMK model (Fig. 3), i.e. a lower temperature change response to cumulative carbon emissions (Fig. 4). Since increasing surface temperature will lead to an additional CO2 release from land to the atmosphere, there is the positive carbon–climate feedback mechanism (Arneth et al., 2010), and here I asked the following question: is there also an effect of the respiration model structure on this feedback strength?
Indeed, this feedback roughly doubles when assuming Michaelis–Menten kinetics, and it is higher for strong carbon emission scenarios (Table 2). As a consequence, the model results imply a higher remaining anthropogenic carbon budget to keep warming below 2 °C above pre-industrial levels of up to circa 100 Pg C but depending on the emission scenario only because we assume an alternative model structure for respiration. These estimates are similar to estimates of additional warming-induced C loss from permafrost-affected soils until 2100 of 10–100 Pg C (Koven et al., 2015). Other additional Earth system feedbacks currently not represented in Earth system models (Sect. 5.5.2.2.5 in Canadell et al., 2023), and additional geophysical uncertainties like non-CO2 forcing or emission uncertainty (Table 5.8 in Canadell et al., 2023), are also of the same order of magnitude. The structural uncertainty in the formulation of respiration is also of the same order of magnitude as the total annual gross primary production or the respiration flux (both 130 Pg C yr−1, Friedlingstein et al., 2023). Shall we assume a linear or non-linear dependence of respiration on the amount of substrate? This assumption influences the internal land carbon dynamics because in the latter case respiration does not respond to higher substrate availability in the same way as in the linear model. This is also visible when looking at the sensitivities of the land carbon change to CO2 change (β, Table 4) which roughly double when assuming Michaelis–Menten kinetics because the response of respiration to higher substrate availability is lower.
I applied a simplified model of global biogeochemical feedback mechanisms, considering only one terrestrial carbon pool, hence integrating autotrophic and heterotrophic respiration and no explicit pool of microbial biomass and microbial functions. Therefore, many specific underlying processes and interactions of ecosystem components are neglected. For example, an increase in heterotrophic respiration due to increasing plant productivity and carbon input to soils (priming effect, Fontaine et al., 2007; Keuper et al., 2020), or changing microbial community structure as a response to climate change (Glassman et al., 2018), is not considered. Nutrient limitation of vegetation productivity (Hungate et al., 2003) is only implicitly parametrized in Eqs. (3) and (4) through a logarithmic response function of GPP to CO2. Hence, I do not quantify the effects of nutrient availability on the carbon–climate feedback in addition to the effects of either respiration model used. When assuming a MMK model, increasing CO2 leads to a higher increase in land C stocks (β, Table 4) due to lower respiration. However, this mechanism can, for instance, also lock more nutrients in soil organic matter and hence change the response function of GPP to CO2. When considering nutrient processes, land C change sensitivities to CO2 and temperature have been shown to be much smaller (Arora et al., 2020). In addition, climate change is expressed as a temperature change in this model, and precipitation effects on carbon cycle functions (Jung et al., 2017) are not taken into account. Therefore, the presented results are first conservative estimates, which should be verified using a state-of-the-art ESM including nutrient cycles and Michaelis–Menten kinetics (Yu et al., 2020).
Besides structural uncertainty, an additional relevant source of uncertainty of such a highly parametrized model is parameter uncertainty. Interestingly, the additional analysis presented in the Supplement shows that the structural uncertainty of the carbon–climate feedback due to the respiration equation is higher than the parameter uncertainty, regardless of the emission scenario applied.
Still, the presented results point to the importance of communicating and addressing existing structural uncertainties in Earth system models. Just assuming an underlying Michaelis–Menten kinetics of respiration processes leads to distinct projections of future respiration and the carbon–climate feedback mechanism. These results also demonstrate the need for novel research, clarifying a valid process-based model structure of ecosystem respiration.
Two major gross carbon fluxes govern the recent land carbon sink, photosynthesis, and respiration. While detailed process-based photosynthesis models have been developed and applied in Earth system models, how to model respiration processes remains unclear. The model structure of respiration alone can lead to a doubling of the carbon–climate feedback estimate over the 21st century. Depending on the underlying emission scenario, that translates into a substantial difference of the remaining carbon budget to keep global warming below 2 °C at up to circa 100 Pg C depending on the emission scenario. These results show the importance of an increased understanding of the mathematical model structure of respiration processes in Earth system models for more reliably projecting future carbon dynamics and climate and related feedback mechanisms and hence estimating a valid remaining anthropogenic carbon budget.
MATLAB code of the model versions applied is available via Zenodo at https://doi.org/10.5281/zenodo.15696851 (Beer, 2025).
Historical CO2 emissions are available at Friedlingstein et al. (2023b). SSP CO2 emission data is publicly available at the SSP database v1.1 (https://tntcat.iiasa.ac.at/SspDb) via the “CMIP6 Emissions” tab.
The supplement related to this article is available online at https://doi.org/10.5194/esd-16-1527-2025-supplement.
The author has declared that there are no competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
Christian Beer acknowledges financial support by the Deutsche Forschungsgemeinschaft through the Heisenberg programme (grant no. 508047523).
This research has been supported by the Deutsche Forschungsgemeinschaft (grant no. 508047523).
This paper was edited by Parvadha Suntharalingam and reviewed by William Wieder and three anonymous referees.
Alexandrov, G. A., Oikawa, T., and Yamagata, Y.: Climate dependence of the CO2 fertilization effect on terrestrial net primary production, Tellus Series B-Chemical and Physical Meteorology, 55, 669–675, https://doi.org/10.1034/j.1600-0889.2003.00021.x, 2003.
Arneth, A., Harrison, S. P., Zaehle, S., Tsigaridis, K., Menon, S., Bartlein, P. J., Feichter, J., Korhola, A., Kulmala, M., O'Donnell, D., Schurgers, G., Sorvari, S., and Vesala, T.: Terrestrial biogeochemical feedbacks in the climate system, Nat. Geosci., 3, 525–532, https://doi.org/10.1038/ngeo905, 2010.
Arora, V. K., Katavouta, A., Williams, R. G., Jones, C. D., Brovkin, V., Friedlingstein, P., Schwinger, J., Bopp, L., Boucher, O., Cadule, P., Chamberlain, M. A., Christian, J. R., Delire, C., Fisher, R. A., Hajima, T., Ilyina, T., Joetzjer, E., Kawamiya, M., Koven, C. D., Krasting, J. P., Law, R. M., Lawrence, D. M., Lenton, A., Lindsay, K., Pongratz, J., Raddatz, T., Séférian, R., Tachiiri, K., Tjiputra, J. F., Wiltshire, A., Wu, T., and Ziehn, T.: Carbon–concentration and carbon–climate feedbacks in CMIP6 models and their comparison to CMIP5 models, Biogeosciences, 17, 4173–4222, https://doi.org/10.5194/bg-17-4173-2020, 2020.
Beer, C.: Simplified model of global biogeochemical feedbacks, Zenodo [code], https://doi.org/10.5281/zenodo.15696851, 2025.
Brovkin, V., Boysen, L., Raddatz, T., Gayler, V., Loew, A., and Claussen, M.: Evaluation of vegetation cover and land-surface albedo in MPI-ESM CMIP5 simulations, Journal of Advances in Modeling Earth Systems, 5, 48–57, https://doi.org/10.1029/2012MS000169, 2013.
Canadell, J. G., Monteiro, P. M. S., Costa, M. H., Cotrim da Cunha, L., Cox, P. M., Eliseev, A. V., Henson, S., Ishii, M., Jaccard, S., Koven, C., Lohila, A., Patra, P. K., Piao, S., Rogelj, J., Syampungani, S., Zaehle, S., and Zickfeld, K.: Global Carbon and Other Biogeochemical Cycles and Feedbacks, in: Climate Change 2021 – The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, edited by: Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge, 673–816, https://doi.org/10.1017/9781009157896.007, 2023.
Cox, P. M., Betts, R. A., Jones, C. D., Spall, S. A., and Totterdell, I. J.: Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model, Nature, 408, 184–187, 2000.
Cramer, W., Bondeau, A., Woodward, F. I., Prentice, I. C., Betts, R. A., Brovkin, V., Cox, P. M., Fisher, V., Foley, J. A., Friend, A. D., Kucharik, C., Lomas, M. R., Ramankutty, N., Sitch, S., Smith, B., White, A., and Young-Molling, C.: Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models, Global Change Biology, 7, 357–373, 2001.
Fontaine, S., Barot, S., Barré, P., Bdioui, N., Mary, B., and Rumpel, C.: Stability of organic carbon in deep soil layers controlled by fresh carbon supply, Nature, 450, 277-U210, https://doi.org/10.1038/nature06275, 2007.
Friedlingstein, P., Dufresne, J. L., Cox, P. M., and Rayner, P.: How positive is the feedback between climate change and the carbon cycle?, Tellus B: Chemical and Physical Meteorology, 55, 692–700, https://doi.org/10.3402/tellusb.v55i2.16765, 2003.
Friedlingstein, P., Cox, P., Betts, R., Bopp, L., Von Bloh, W., Brovkin, V., Cadule, P., Doney, S., Eby, M., Fung, I., Bala, G., John, J., Jones, C., Joos, F., Kato, T., Kawamiya, M., Knorr, W., Lindsay, K., Matthews, H. D., Raddatz, T., Rayner, P., Reick, C., Roeckner, E., Schnitzler, K. G., Schnur, R., Strassmann, K., Weaver, A. J., Yoshikawa, C., and Zeng, N.: Climate-carbon cycle feedback analysis: Results from the C4MIP model intercomparison, J. Climate, 19, 3337–3353, 2006.
Friedlingstein, P., O'Sullivan, M., Jones, M. W., Andrew, R. M., Bakker, D. C. E., Hauck, J., Landschützer, P., Le Quéré, C., Luijkx, I. T., Peters, G. P., Peters, W., Pongratz, J., Schwingshackl, C., Sitch, S., Canadell, J. G., Ciais, P., Jackson, R. B., Alin, S. R., Anthoni, P., Barbero, L., Bates, N. R., Becker, M., Bellouin, N., Decharme, B., Bopp, L., Brasika, I. B. M., Cadule, P., Chamberlain, M. A., Chandra, N., Chau, T.-T.-T., Chevallier, F., Chini, L. P., Cronin, M., Dou, X., Enyo, K., Evans, W., Falk, S., Feely, R. A., Feng, L., Ford, D. J., Gasser, T., Ghattas, J., Gkritzalis, T., Grassi, G., Gregor, L., Gruber, N., Gürses, Ö., Harris, I., Hefner, M., Heinke, J., Houghton, R. A., Hurtt, G. C., Iida, Y., Ilyina, T., Jacobson, A. R., Jain, A., Jarníková, T., Jersild, A., Jiang, F., Jin, Z., Joos, F., Kato, E., Keeling, R. F., Kennedy, D., Klein Goldewijk, K., Knauer, J., Korsbakken, J. I., Körtzinger, A., Lan, X., Lefèvre, N., Li, H., Liu, J., Liu, Z., Ma, L., Marland, G., Mayot, N., McGuire, P. C., McKinley, G. A., Meyer, G., Morgan, E. J., Munro, D. R., Nakaoka, S.-I., Niwa, Y., O'Brien, K. M., Olsen, A., Omar, A. M., Ono, T., Paulsen, M., Pierrot, D., Pocock, K., Poulter, B., Powis, C. M., Rehder, G., Resplandy, L., Robertson, E., Rödenbeck, C., Rosan, T. M., Schwinger, J., Séférian, R., Smallman, T. L., Smith, S. M., Sospedra-Alfonso, R., Sun, Q., Sutton, A. J., Sweeney, C., Takao, S., Tans, P. P., Tian, H., Tilbrook, B., Tsujino, H., Tubiello, F., van der Werf, G. R., van Ooijen, E., Wanninkhof, R., Watanabe, M., Wimart-Rousseau, C., Yang, D., Yang, X., Yuan, W., Yue, X., Zaehle, S., Zeng, J., and Zheng, B.: Global Carbon Budget 2023, Earth Syst. Sci. Data, 15, 5301–5369, https://doi.org/10.5194/essd-15-5301-2023, 2023.
Friedlingstein, P., O'Sullivan, M., Jones, M. W., Andrew, R. M., Bakker, D. E. C., Hauck, J., Landschützer, P., Le Quéré, C., Luijkx, I. T., Peters, G. P., Peters, W., Pongratz, J., Schwingshackl, C, Sitch, S., Canadell, J. G., Ciais, P., Jackson, R. B., Alin, S. R., Anthoni, P., Barbero, L., Bates, N. R., Becker, M., Bellouin, N., Decharme, B., Bopp, L., Brasika, I. B. M., Cadule, P., Chamberlain, M. A., Chandra, N., Chau, T.-T.-T., Chevallier, F., Chini, L. P., Cronin, M., Dou, X., Enyo, K., Evans, W., Falk, S., Feely, R. A., Feng, L., Ford, D. J., Gasser, T., Ghattas, J., Gkritzalis, T., Grassi, G., Gregor, L., Gruber, N., Gürses, Ö., Harris, I., Hefner, M., Heinke, J. Houghton, R. A., Hurtt, G. C., Iida, Y., Ilyina, T., Jacobson, A. R., Jain, A., Jarníková, T., Jersild, A., Jiang, F., Jin, Z., Joos, F., Kato, E., Keeling, R. F., Kennedy, K., Goldewijk, K. K., Knauer, J., Korsbakken, J. I., Körtzinger, A., Lan, X., Lefèvre, N., Li, H., Liu, J., Liu, Z., Ma, L., Marland, G., Mayot, N., McGuire, P. C., McKinley, G. A., Meyer, G., Morgan, E. J., Munro, D. R., Nakaoka, S.-I., Niwa, Y., O’Brien, K. M., Olsen, A., Omar, A. M., Ono, T., Paulsen, M., Pierrot, D., Pocock, K., Poulter, B., Powis, C. M., Rehder, G., Resplandy, L., Robertson, E., Rödenbeck, C., Rosan, T. M., Schwinger, J., Séférian, R., Smallman, T. L., Smith, S. M., Sospedra-Alfonso, R., Sun, Q., Sutton, A. J., Sweeney, C., Takao, S., Tans, P. P., Tian, H., Tilbrook, B., Tsujino, H., Tubiello, F., van der Werf, G. R., van Ooijen, E., Wanninkhof, R., Watanabe, M., Wimart-Rousseau, C., Yang, D., Yang, X., Yuan, W., Yue, X., Zaehle, S., Zeng, K., and Zheng, B.: Supplemental data of the Global Carbon Budget 2023, ICOS-ERIC Carbon Portal [data set], https://doi.org/10.18160/GCP-2023, 2023.
Gidden, M. J., Riahi, K., Smith, S. J., Fujimori, S., Luderer, G., Kriegler, E., van Vuuren, D. P., van den Berg, M., Feng, L., Klein, D., Calvin, K., Doelman, J. C., Frank, S., Fricko, O., Harmsen, M., Hasegawa, T., Havlik, P., Hilaire, J., Hoesly, R., Horing, J., Popp, A., Stehfest, E., and Takahashi, K.: Global emissions pathways under different socioeconomic scenarios for use in CMIP6: a dataset of harmonized emissions trajectories through the end of the century, Geosci. Model Dev., 12, 1443–1475, https://doi.org/10.5194/gmd-12-1443-2019, 2019.
Glassman, S. I., Weihe, C., Li, J. H., Albright, M. B. N., Looby, C. I., Martiny, A. C., Treseder, K. K., Allison, S. D., and Martiny, J. B. H.: Decomposition responses to climate depend on microbial community composition, P. Natl. Acad. Sci. USA, 115, 11994–11999, https://doi.org/10.1073/pnas.1811269115, 2018.
Hansen, J., Lacis, A., Rind, D., Russell, G., Stone, P., Fung, I., Ruedy, R., and Lerner, J.: Climate Sensitivity: Analysis of Feedback Mechanisms, in: Climate Processes and Climate Sensitivity, 130–163, https://doi.org/10.1029/GM029p0130, 1984.
Heinze, C., Eyring, V., Friedlingstein, P., Jones, C., Balkanski, Y., Collins, W., Fichefet, T., Gao, S., Hall, A., Ivanova, D., Knorr, W., Knutti, R., Löw, A., Ponater, M., Schultz, M. G., Schulz, M., Siebesma, P., Teixeira, J., Tselioudis, G., and Vancoppenolle, M.: ESD Reviews: Climate feedbacks in the Earth system and prospects for their evaluation, Earth Syst. Dynam., 10, 379–452, https://doi.org/10.5194/esd-10-379-2019, 2019.
Hugelius, G., Strauss, J., Zubrzycki, S., Harden, J. W., Schuur, E. A. G., Ping, C.-L., Schirrmeister, L., Grosse, G., Michaelson, G. J., Koven, C. D., O'Donnell, J. A., Elberling, B., Mishra, U., Camill, P., Yu, Z., Palmtag, J., and Kuhry, P.: Estimated stocks of circumpolar permafrost carbon with quantified uncertainty ranges and identified data gaps, Biogeosciences, 11, 6573–6593, https://doi.org/10.5194/bg-11-6573-2014, 2014.
Hungate, B. A., Dukes, J. S., Shaw, M. R., Luo, Y. Q., and Field, C. B.: Nitrogen and climate change, Science, 302, 1512–1513, https://doi.org/10.1126/science.1091390, 2003.
Jones, C. D., Ziehn, T., Anand, J., Bastos, A., Burke, E., Canadell, J. G., Cardoso, M., Ernst, Y., Jain, A. K., Jeong, S., Keller, E. D., Kondo, M., Lauerwald, R., Lin, T.-S., Murray-Tortarolo, G., Nabuurs, G.-J., O'Sullivan, M., Poulter, B., Qin, X., von Randow, C., Sanches, M., Schepaschenko, D., Shvidenko, A., Smallman, T. L., Tian, H., Villalobos, Y., Wang, X., and Yun, J.: RECCAP2 Future Component: Consistency and Potential for Regional Assessment to Constrain Global Projections, AGU Adv., 4, e2023AV001024, https://doi.org/10.1029/2023AV001024, 2023.
Jung, M., Reichstein, M., Schwalm, C. R., Huntingford, C., Sitch, S., Ahlström, A., Arneth, A., Camps-Valls, G., Ciais, P., Friedlingstein, P., Gans, F., Ichii, K., Ain, A. K. J., Kato, E., Papale, D., Poulter, B., Raduly, B., Rödenbeck, C., Tramontana, G., Viovy, N., Wang, Y. P., Weber, U., Zaehle, S., and Zeng, N.: Compensatory water effects link yearly global land CO sink changes to temperature, Nature, 541, 516–520, https://doi.org/10.1038/nature20780, 2017.
Keuper, F., Wild, B., Kummu, M., Beer, C., Blume-Werry, G., Fontaine, S., Gavazov, K., Gentsch, N., Guggenberger, G., Hugelius, G., Jalava, M., Koven, C., Krab, E. J., Kuhry, P., Monteux, S., Richter, A., Shahzad, T., Weedon, J. T., and Dorrepaal, E.: Carbon loss from northern circumpolar permafrost soils amplified by rhizosphere priming, Nat. Geosci., 13, 560–565, https://doi.org/10.1038/s41561-020-0607-0, 2020.
Koven, C. D., Schuur, E. A. G., Schädel, C., Bohn, T. J., Burke, E. J., Chen, G., Chen, X., Ciais, P., Grosse, G., Harden, J. W., Hayes, D. J., Hugelius, G., Jafarov, E. E., Krinner, G., Kuhry, P., Lawrence, D. M., MacDougall, A. H., Marchenko, S. S., McGuire, A. D., Natali, S. M., Nicolsky, D. J., Olefeldt, D., Peng, S., Romanovsky, V. E., Schaefer, K. M., Strauss, J., Treat, C. C., and Turetsky, M.: A simplified, data-constrained approach to estimate the permafrost carbon-climate feedback, Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 373, 20140423, https://doi.org/10.1098/rsta.2014.0423, 2015.
Koven, C. D., Sanderson, B. M., and Swann, A. L. S.: Much of zero emissions commitment occurs before reaching net zero emissions, Environ. Res. Lett., 18, 014017, https://doi.org/10.1088/1748-9326/acab1a, 2023.
Lade, S. J., Donges, J. F., Fetzer, I., Anderies, J. M., Beer, C., Cornell, S. E., Gasser, T., Norberg, J., Richardson, K., Rockström, J., and Steffen, W.: Analytically tractable climate–carbon cycle feedbacks under 21st century anthropogenic forcing, Earth Syst. Dynam., 9, 507–523, https://doi.org/10.5194/esd-9-507-2018, 2018.
Lloyd, J. and Taylor, J. A.: On the temperature dependence of soil respiration, Functional Ecology, 8, 315–323, 1994.
Nijsse, F. J. M. M., Cox, P. M., and Williamson, M. S.: Emergent constraints on transient climate response (TCR) and equilibrium climate sensitivity (ECS) from historical warming in CMIP5 and CMIP6 models, Earth Syst. Dynam., 11, 737–750, https://doi.org/10.5194/esd-11-737-2020, 2020.
O'Sullivan, M., Friedlingstein, P., Sitch, S., Anthoni, P., Arneth, A., Arora, V. K., Bastrikov, V., Delire, C., Goll, D. S., Jain, A., Kato, E., Kennedy, D., Knauer, J., Lienert, S., Lombardozzi, D., McGuire, P. C., Melton, J. R., Nabel, J. E. M. S., Pongratz, J., Poulter, B., Séférian, R., Tian, H. Q., Vuichard, N., Walker, A. P., Yuan, W. P., Yue, X., and Zaehle, S.: Process-oriented analysis of dominant sources of uncertainty in the land carbon sink, Nat. Commun., 13, 4781, https://doi.org/10.1038/s41467-022-32416-8, 2022.
Reichstein, M. and Beer, C.: Soil respiration across scales: the importance of a model–data integration framework for data interpretation, Journal of Plant Nutrition and Soil Science, 171, 344–354, 2008.
Riahi, K., van Vuuren, D. P., Kriegler, E., Edmonds, J., O'Neill, B. C., Fujimori, S., Bauer, N., Calvin, K., Dellink, R., Fricko, O., Lutz, W., Popp, A., Cuaresma, J. C., Samir, K. C., Leimbach, M., Jiang, L., Kram, T., Rao, S., Emmerling, J., Ebi, K., Hasegawa, T., Havlik, P., Humpenöder, F., Silva, L. A. D., Smith, S., Stehfest, E., Bosetti, V., Eom, J., Gernaat, D., Masui, T., Rogelj, J., Strefler, J., Drouet, L., Krey, V., Luderer, G., Harmsen, M., Takahashi, K., Baumstark, L., Doelman, J. C., Kainuma, M., Klimont, Z., Marangoni, G., Lotze-Campen, H., Obersteiner, M., Tabeau, A., and Tavoni, M.: The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview, Global Environmental Change, 42, 153–168, 2017.
Sitch, S., Smith, B., Prentice, I. C., Arneth, A., Bondeau, A., Cramer, W., Kaplan, J. O., Levis, S., Lucht, W., Sykes, M. T., Thonicke, K., and Venevsky, S.: Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model, Global Change Biology, 9, 161–185, https://doi.org/10.1046/j.1365-2486.2003.00569.x, 2003.
Tang, J., Riley, W. J., and Zhu, Q.: Supporting hierarchical soil biogeochemical modeling: version 2 of the Biogeochemical Transport and Reaction model (BeTR-v2), Geosci. Model Dev., 15, 1619–1632, https://doi.org/10.5194/gmd-15-1619-2022, 2022.
Vaughn, L. J. S. and Torn, M. S.: C evidence that millennial and fast-cycling soil carbon are equally sensitive to warming, Nature Climate Change, 9, 467–471, https://doi.org/10.1038/s41558-019-0468-y, 2019.
Wieder, W. R., Bonan, G. B., and Allison, S. D.: Global soil carbon projections are improved by modelling microbial processes, Nature Climate Change, 3, 909–912, https://doi.org/10.1038/nclimate1951, 2013.
Yu, L., Ahrens, B., Wutzler, T., Schrumpf, M., and Zaehle, S.: Jena Soil Model (JSM v1.0; revision 1934): a microbial soil organic carbon model integrated with nitrogen and phosphorus processes, Geosci. Model Dev., 13, 783–803, https://doi.org/10.5194/gmd-13-783-2020, 2020.
Zickfeld, K., Eby, M., Matthews, H. D., Schmittner, A., and Weaver, A. J.: Nonlinearity of Carbon Cycle Feedbacks, J. Climate, 24, 4255–4275, https://doi.org/10.1175/2011JCLI3898.1, 2011.